Introduction
Alveoli represent the most distal portion of the respiratory tract and are major sites of gas exchange. Each alveolus is covered with pulmonary capillaries for gas exchange and is composed of type 1 and 2 alveolar epithelial cells (AECs) (hereafter called AT1 and AT2 cells, respectively) and macrophages. AT1 cells are thin, squamous epithelial cells that cover approximately 80% of the alveolar surface area and are connected with other AECs by tight junctions, forming an impermeable barrier. In contrast, AT2 cells cover approximately only 5% of the alveolar surface. Despite their small number, AT2 cells play pivotal roles in retaining alveolar function and integrity, including (1) synthesis and secretion of surfactant, (2) host defense, (3) fluid balance, and (4) regeneration of the alveolar epithelium following injury [1]. Alveolar macrophages (AMs) reside in the alveoli, where they play key roles as the first line of defense in scavenging various pathogens and process inhaled environmental particles [2,3]. Thus, immunological functions of AMs are implicated in the pathological development of acute and chronic pulmonary diseases.
Pulmonary diseases such as idiopathic pulmonary fibrosis (IPF) and chronic obstructive pulmonary disease (COPD) result in alveolar injury. The damage evoked in the alveolar epithelial tissue of IPF and COPD patients is irreversible and eventually leads to death. Moreover, AT2 cells are among the primary target cells for severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection, as they express angiotensin-converting enzyme 2 (ACE2), a receptor of the virus and a main entry point for SARS-CoV-2 infection [4]. Once AT2 cells are damaged by viral infection, subsequent vascular leakage triggers a hyperinflammatory state, resulting in severe respiratory complications. Thus, AT2 cells could be a promising therapeutic target for the treatment of both chronic pulmonary diseases such as IPF and COPD and fatal coronavirus infections. To understand the underlying pathogenesis of these fatal acute and chronic respiratory diseases, techniques have been developed to isolate and purify AT1 and AT2 cells from human lung tissue. Human primary AECs more closely represent the in vivo state of alveoli and are expected to yield the most biologically accurate outcomes [5]. However, their low accessibility, limited proliferative lifespan in culture, and loss of functional properties over time remain bottlenecks for their use. Furthermore, two-dimensional AEC cultures are not entirely representative for the whole alveolar tissue due to low cellular heterogeneity and architectural similarity. In order to overcome limited lifespan, a genetically modified pulmonary epithelial cell line (A549) has been widely used to model respiratory diseases for biopharmaceutical research and to evaluate the pulmotoxicity of suspected harmful materials. However, these cells exhibit phenotypic and functional differences and respond to toxins differently compared to the in vivo alveolar tissue. Alternatively, primary AECs from laboratory animals can be used, but species-specific differences in the molecular mechanisms of pulmonary diseases and responses to viral infection may also limit their applicability to humans. For example, mouse AT2 cells form functional monolayers on laminin 5 extracellular matrix (ECM) to achieve tight barrier properties that actively absorb sodium ion in contrast to their human counterparts [6]. Moreover, transgenic mice expressing human ACE2 should be used for the study of SARS-CoV-2 infection due to absence of ACE2 in AT2 of inbred wild-type mice [7]. Thus, there is a profound need to develop human-relevant alveolar models from renewable biological sources that can give rise to AECs that better resemble human primary AECs at the phenotypic, genetic, and functional levels.
Human pluripotent stem cells (hPSCs) have demonstrated great value as an alternative source that can give rise to all somatic cell types, including airway and AECs. Thus, hPSCs provide a unique opportunity to generate three-dimensional (3D) configurations, such as spheroids and organoids, that recapitulate the complexity and functions of in vivo tissues. Three types of cells are typically used for lung alveolar epithelial differentiation and generation of alveolar organoids (AOs); human embryonic stem cells (hESCs), human-induced pluripotent stem cells (hiPSCs) and adult or fetal tissue. Tissue-derived AOs are the most ideal AOs for development and disease study among the three types because the tissue sources were adapted to a lung tissue-specific niche for a long time. However, since tissue-derived AOs have donor-dependent characteristics, it is difficult to confirm consistent reactivity and still have the limitation of being heterogeneous organoids in terms of size and morphology. On the other side, hESCs and hiPSCs do not have problem of donor dependence, but have the weakness of immune rejection from a therapeutic perspective. In particular, hESCs have the limitation that they are always accompanied by ethical issues [8]. Nevertheless, in recent years hPSCs have been successfully employed to generate several types of organoids representing different respiratory compartments, including alveolar regions [9-12]. More recently, hPSC-derived AOs with genetic defects associated with cystic fibrosis, surfactant deficiency, and impaired surfactant secretion have been generated to support studies to understand the roles of specific cell types during pathological development [13-15]. Despite continued advances in 3D culture techniques and single-cell genomics, there is still a profound need to improve the cellular heterogeneity and maturity of AOs to recapitulate key histological and functional features of in vivo alveolar tissue. In particular, the incorporation of immune cells such as macrophages into hPSC-AO systems is crucial for disease modeling and subsequent drug screening. In this review, we summarize current methods for differentiating AT2 cells from hPSCs followed by AO generation and their applications in disease modeling, drug testing, and toxicity evaluation. Furthermore, we describe how current hPSC-derived AOs closely resemble in vivo alveoli in terms of phenotype, cellular heterogeneity, and maturity.
Definitive Endoderm Differentiation
The endoderm is the innermost germ layer of that generates the respiratory organs (lung and trachea), the gastrointestinal tract (gut, liver, and pancreas), and the thyroid [16]. Lung is the vital organ and susceptible to life-threatening diseases, especially fatal virus infection. Thus, the production of high-quality endodermal cells on a large scale provides a unique opportunity to model acute and chronic respiratory diseases and apply them for translational study [17]. In all current protocols, the induction of undifferentiated hPSCs into definitive endoderm (DE) is the first step for their further differentiation into functional and mature AECs. The guided differentiation of hPSCs into DE is induced with a high dose of activin A (100 ng/mL), which mimics the action of Nodal, a protein that signals via activation of its effector suppressor mothers against decapentaplegic homolog 2/3 (SMAD2/3) to promote endodermal differentiation during embryonic gastrulation [16]. Additionally, a wingless-related integration site (Wnt) is required for the normal formation of the primitive streak [18] and the induction of the key endoderm marker SRY-related high-mobility group box 17 (SOX17) [18,19]. Therefore, these protocols have been developed in combination with CHIR99021 (CHIR), a Wnt activator, to enhance DE differentiation. Consequently, the efficiency of DE differentiation can be easily determined by flow cytometry analysis of the frequencies of DE markers, such as C-X-C chemokine receptor type 4 (CXCR4), SOX17, GATA-binding protein 6 (GATA6), and forkhead box protein A2 (FOXA2).
However, homogeneous and reproducible differentiation still remains a challenge, highlighting the need for further understanding of the molecular mechanisms regulating DE differentiation based on fundamental principles learned from early development. Chia et al. [20] showed that inhibition of phosphatidylinositol 3-kinase/rapamycin complex 2 reduces the phosphorylation of the SMAD2/3-T220/T179 linker residue and increases the duration of SMAD2/3 activity, promoting a more robust endoderm differentiation of hESCs. During gastrulation, GATA6 is expressed in the DE, suggesting that understanding the mechanisms regulating GATA6 expression may contribute to the efficient differentiation of DE from hPSCs. Interestingly, GATA6 heterozygous hPSCs show a modest reduction in DE formation, while GATA6-null hPSCs fail to enter the DE lineage [17]. During DE differentiation, GATA6 binds to and cooperates with eomesodermin and SMAD2/3 to regulate the endodermal lineage specification. Furthermore, recent transcriptomic analysis has revealed that GATA6-antisense RNA1 (AS1), a long noncoding RNA divergently transcribed from the GATA6 locus, is highly expressed during DE differentiation and interacts with SMAD2/3 to activate GATA6 [21]. These results indicate that GATA6 plays a key role as a gatekeeper to early DE differentiation. Recent genome-scale clustered regularly interspaced short palindromic repeats (CRISPR) screens identified JUN N-terminal kinase (JNK)-JUN signaling as a key barrier to DE differentiation. The screens showed that pharmacological inhibition of JNK significantly improves the efficiencies of generating DE and DE-derived lung progenitors by reconfiguring SMAD2/3 binding to enhancers cobound by GATA6 [22]. More interestingly, JNK inhibition reduces the dose requirements for activin A (20 ng/mL instead of the typical 100 ng/mL) for efficient DE differentiation. These studies highlight the potential of harnessing the knowledge from studies of early embryonic development for homogeneous and efficient DE differentiation.
Heterogeneity and Plasticity in hPSC-Derived AT2 Cells
As described above, despite their small number, AT2 cells play a pivotal role in retaining alveolar function and repairing damage through self-renewal and differentiation into AT1 cells [23,24]. Thus, identification of alveolar epithelial progenitors (AEPs) and their subsequent maturation towards functional AT2 cells during hPSC differentiation are of great interest for generating functional AOs to improve the understanding of alveolar development, regeneration, and pathogenesis. NK2 homeobox 1 (NKX2.1) is a transcription factor and is utilized as a key marker gene to determine the differentiation of hPSCs towards lung epithelial progenitors. Using an NKX2.1 knock-in reporter hPSC line, NKX2.1+ AEPs (CD47hi/CD26lo) can be isolated by fluorescence-activated cell sorting (FACS) and further utilized to generate renewable surfactant-associated protein C-positive (SFTPC+) AT2 cells as epithelial-only spheres for up to 1 year [25]. Alternatively, carboxypeptidase M (CPM) has been newly identified as a surface marker of NKX2-1+ ventralized anterior foregut endoderm cells (VAFECs) in vitro and in fetal human and murine lungs [25,26]. As reported in these two studies, FACS-based isolation of CPM+ cells allowed the generation of long-term expandable AOs harboring self-renewing SFTPC+ AT2 cells that can differentiate into AT1 cells.
One of the key roles of AT2 cells is to synthesize and secrete pulmonary surfactant for the maintenance of alveolar surface tension [27]. Thus, most studies have employed SFTPC as a key marker distinguishing mature and functional AT2 cells from immature and non-AT2 cells during stepwise AEC differentiation from pluripotent stem cells in mouse and human tissues [25,26,28,29]. Interestingly, recent single-cell RNA sequencing profiling revealed a gradient expression of SFTPC differentiating less mature embryonic lung progenitors and more mature AT2 cells [30]. Furthermore, AT2 cells can be also distinguished from more proliferative and highly Wnt-responsive subpopulations from less-responsive ones in murine lung tissues [31-33]. Similarly, heterogeneity in human AT2 cells has been described using reporter cell lines. Sun et al. [29] established an ATP binding cassette subfamily A member 3 (ABCA3):green fluorescent protein (GFP) and SFTPCtdTomato bifluorescent reporter hPSC line and compared ABCA3:GFP/SFTPCtdTomato double-positive (DP) with ABCA3:GFP single-positive (SP) subsets by RNA sequencing and functional assays and found that the DP and SP subsets were distinct in terms of AT2 cell function, transcriptomic maturity, proliferative activity, clonogenicity, and surfactant secretion capability. More interestingly, each population gave rise to the other, suggesting some interchangeability [29]. These findings suggest that AT2 cells are capable of some heterogeneity and plasticity, which may make it difficult to understand how AT2 cells contribute to lung homeostasis and repair alveolar damage.
The reason heterogeneity exists in AT2 cells is for maintaining homeostasis of lung tissue and for effective regeneration upon injury [34]. In the steady-state condition, quiescent cells are suitable for maintaining homeostasis, while rapid conversion to cells suitable for division and differentiation is required in case of injury. Indeed, various subpopulations of AT2 cells have been identified in mouse models both at normal and under injury conditions and subtypes of AT2 cells in both conditions exhibited different characteristics. Additionally, heterogeneous AT2 subpopulations have been identified in the hPSC-derived AT2 cells [29]. However, the presence and function of heterologous AT2 cell subpopulations similar to those above have not yet been identified in human lung tissue. Thus, achieving a deeper understanding of the heterogeneity in AT2 cells is important for deriving functional and mature AOs resembling human adult alveoli for disease modeling, drug testing, and other applications. Interestingly, beyond the notion that AT2 cells act like lung stem cells, differentiating into AT1 cells to regulate lung homeostasis and respond to injury, it has recently been reported that AT1 cells also have the plasticity to differentiate into other cells [35]. Shen et al. [35] identified a new lung-specific AT1 cell marker, GRAM domain containing 2 (Gramd2) by generating Gramd2CreERT2 transgenic strain and confirmed that Gramd2CreERT2 GFP+ cells were able to generate the organoids when co-cultured with mouse fibroblasts. Intriguingly, even though the organoids were created by GFP+ AT1 cells, other cell types such as club cells, basal cells, and AT2 cells were identified over time. Gramd2+ AT1 cells were able to transition through various intermediate lung epithelial cell states such as basal, secretory and AT2 cells in organoids while acquiring proliferative capacity. These findings indicate that Gramd2+ AT1 cells are highly plastic, suggesting AT1 cells also may contribute to alveolar regeneration upon injury. Therefore, establishing a lung disease model based on hPSC-derived AOs will be very useful to clarify the role of heterogeneous epithelial cells and their repair and regeneration mechanisms.
Generation of hPSC-Derived AOs and Their Applications
The current methods for generating hPSC-derived AOs are based on the Gotoh method, which was first reported in the generation of pulmonary AOs [26]. On day 14 of alveolar differentiation, progenitor stage NKX2.1+ cells are isolated from the CD47hi/CD26lo population using FACS and then embedded in Matrigel. Ten days after Matrigel embedding, a rounder organoid structure is generated that is capable of expressing alveolar epithelial genes, such as SFTPC, surfactant-associated protein B (SFTPB), surfactant-associated protein A (SFTPA), and aquaporin 5 (AQP5). Moreover, derived AOs are composed of three types of cells, namely, AT1, AT2, and progenitor cells, and have lamellar body-like structures that are important for surfactant protein storage and secretion. These organoids are capable of long-term expansion up to 119 days without genetic or structural alterations and are applicable to pulmonary disease modeling and drug toxicology studies recapitulating AT2-cell-specific phenotypes [36].
In the formation of hPSC-derived AOs, different research strategies have been utilized for fibroblast-dependent and fibroblast-free conditions. Mesenchymal cells are essential to lung development and are known to promote AEC differentiation and homeostasis through epithelial-mesenchymal interactions. In fact, mixing mesenchymal cells with fibroblast cells followed by treatment with fibroblast-conditioned media can significantly facilitate the generation of AOs. Gotoh et al. [26] established a 3D co-culture system with fetal human fibroblasts to promote organoid formation, and Barkauskas et al. [23] confirmed that more organoids were formed in the fibroblast-dependent condition using primary platelet-derived growth factor receptor ╬▒-positive (PDGFR╬▒+) lung fibroblasts. Meanwhile, Yamamoto et al. [36] optimized a 3D AO protocol that did not require the support of fibroblast cells. The fibroblast-free condition did not affect the differentiation and proliferation of hPSC-derived AT2 cells and successfully allowed the generation of fibroblast-free AOs harboring SFTPC+ cells with lamellar bodies. The fibroblast-free condition facilitates the generation of AOs composed of pure AT2 cells. However, many reports still indicate that fibroblast-dependent conditions favor the induction and stable expansion of the SFTPC + cell population through epithelial-mesenchymal interactions. As the incorporation of fibroblast cells in the formation of AOs is still controversial, the need for fibroblasts in these applications requires further investigation.
hPSC-derived AOs have contributed to a deeper understating of pulmonary pathogenesis by providing useful platforms for disease modeling and drug testing (Table 1) [37-39]. Treatment with transforming growth factor-╬▓1 (TGF-╬▓1) or bleomycin-induced fibrotic changes in hPSC-derived AOs through the accumulation of ECM and the upregulation of epithelial-to-mesenchymal transition-related genes [40]. Using this fibrosis modeling strategy, the efficacies and pharmacological mechanisms of action of various potential drugs have been investigated. Kim et al. [40] showed that the fibrotic response of TGF-╬▓1-induced AOs was significantly diminished following treatment with NP-011 (a truncated form of the milk fat globule epidermal growth factor 8 [MFG-E8] protein) through a mechanism involving the suppression of extracellular signal-related kinase signaling. In addition, the fibrosis response was alleviated by inhibiting activin receptor-like kinase 5 and blocking integrin ╬▒v╬▓6, and the effects of the fibrosis clinical drugs nintedanib and pirfenidone were evaluated using hPSC-derived AOs, experiments that confirmed that epithelial reprogramming was not completely reversed, whereas ECM deposition and pro-fibrotic factors were reduced [38]. In addition to the fibrosis model, AOs have been actively applied to assess environmental pollutant toxicity. Exposure of hPSC-derived AOs to fine particulate matter 2.5 (PM2.5) revealed a significant decrease in AO diameter compared with control cells and induced inflammatory and fibrotic responses [41]. Interestingly, treatment with diesel fine PM2.5 (dPM2.5) significantly increased the expression levels of ACE2 and transmembrane protease serine 2, which are key receptors mediating SARS-CoV-2 entry into host cells. More recently, the cytotoxicities of nine different chemicals classified as ŌĆ£acute toxicity, inhalationŌĆØ hazards were assessed in a 3D AO culture system, indicating that AO-based 3D culture systems provide a useful platform for modeling chemical exposures [42].
Additionally, hPSC-derived AOs can be applied to more disease-specific pathogenesis studies by generating disease-specific genetic defects in hPSCs and using patient-derived stem cells. Strikoudis et al. [14] generated some gene-deficient hPSC line associated with Hermansky-Pudlak syndrome (HPS) to generate HPS-associated interstitial pneumonia (HPSIP) specific AO models. Depletion of HPS1, HPS2, and HPS4 genes increased interleukin-11 levels in hPSC-derived epithelial cells and promoted spontaneous lung fibrosis in lung organoids. In addition, its potential as a therapeutic target for pulmonary fibrosis was demonstrated by confirming that pulmonary fibrosis was alleviated through deletion of interleukin 11 [14]. To implement severe phenotypes of HPSIP in AOs, other researcher generated patient-specific induced pluripotent stem cell (iPSCs) from patient-derived fibroblasts with the most common bi-allelic variant, c.1472_1487dup16, in HPS1 [43]. The lung bud organoids (LBOs) derived from patient-specific iPSCs successfully implemented the abnormalities in morphology and size that is representative characteristic of HPS1. Mitochondrial dysfunction in HPS1 patient-specific AECs were confirmed via proteomic analysis of AOs and giant lamellar bodies that is another feature of HPS1 were observed in iPSC-derived AT2 cells. In another paper, SFTPB-deficient (p.Pro133GlnfsTer95) hiPSCs were generated from patient-specific fibroblasts to closely represent the lungs of human fetuses with fatal respiratory distress secondary to SFTPB deficiency. Using this disease-specific AO model, researcher demonstrated that lentiviral correction therapy is applicable for fetal disease caused by SFTPB deficiency in vitro [13]. More recently, using patient-specific iPSCs carrying an AEC2-exclusive disease-related variant (SFTPCI73T), a human in vitro model demonstrating epithelial-intrinsic events resulting in AEC2 dysfunction has been reported [44]. AT2 cells derived from the I73T mutant cell line showed similar changes to in vivo alteration such as accumulation of large amounts of misprocessed and misimported pro-SFTPC proteins.
hPSC-AOs: Cellular Heterogeneity
hPSC-derived AOs have facilitated the study of pulmonary disease modeling, drug testing, and toxic material assessment by mimicking the physiology and pathophysiology of human lung tissue [45-47]. Despite the successful production of AOs and their application in pulmonary disease modeling and drug screening studies, more efforts such as improving their cellular diversity and maturity will be required to recapitulate the complex human alveolar environment. The environment and function of human lung alveoli are regulated through interactions between various cells that comprise the lung, such as AT1 and AT2 epithelial cells, fibroblasts, macrophages, and vascular cells. Achieving a detailed understanding of how these cell types interact with each other in the pathogenesis of pulmonary diseases is fundamental for designing control and treatment strategies. Therefore, it is necessary to establish advanced 3D multicellular AO culture systems containing key alveolar components.
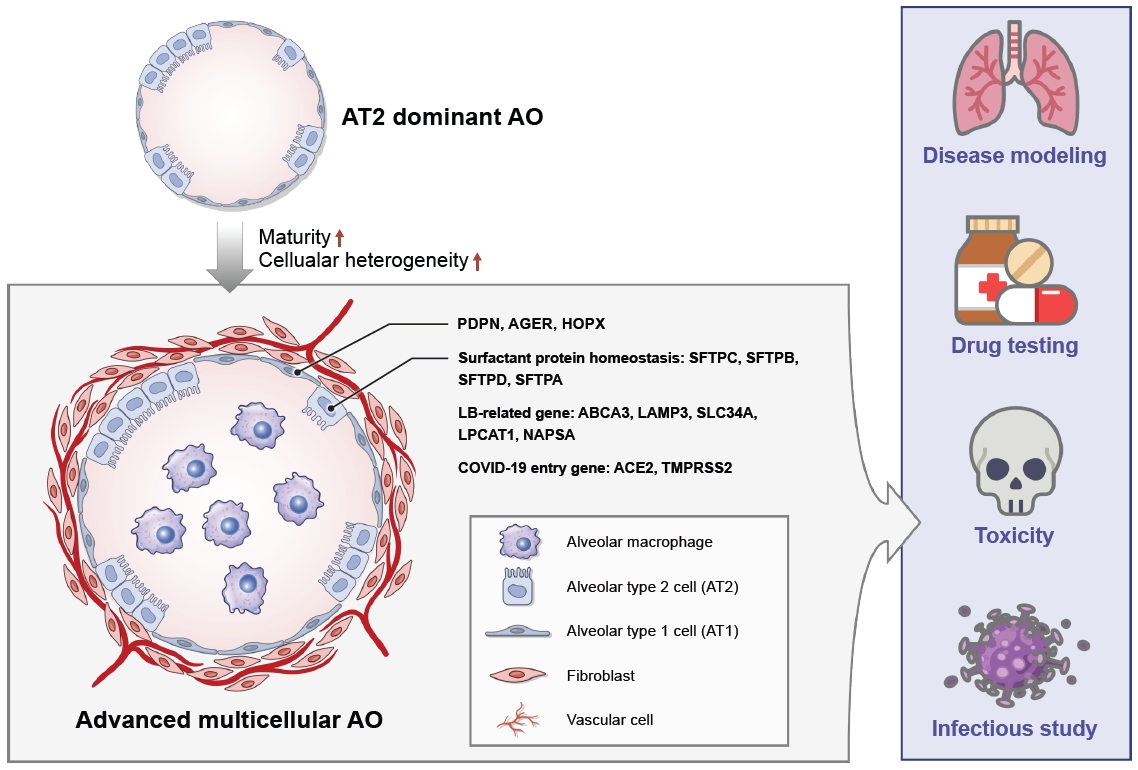
Macrophages are crucial to the immune defense system and are closely associated with pulmonary fibrosis, acting as a double-edged sword to either trigger or suppress disease depending on their polarization [48]. Thus, the incorporation of macrophages into hPSC-AO systems is crucial for disease modeling and subsequent drug screening (Figure 1). Heo and Hong [49] first established a multicellular alveolar organoid (Mac-AO) system containing functional macrophages derived from hPSCs. The macrophages were aggregated with AECs at a 1:5 ratio and cultured for 1 week under AO media conditions. In the Mac-AOs, the macrophage phenotype and number were well maintained without phenotypic alteration. Importantly, Mac-AOs showed more potential for fibrosis modeling and drug efficacy testing applications due to their strong fibrotic response to TGF-╬▓ treatments. Seo et al. [50] generated Mac-AOs that mimic the structure of human alveoli in which the macrophages are located within the alveoli by direct injection of hPSC-derived macrophages into the AOs using a syringe. During the 2-week culture of Mac-AOs, changes in the macrophage secretome in response to lipopolysaccharide inflammation were identified, demonstrating a possible secretory function of macrophages in AOs.
In the case of brain organoids, studies on immune cell-containing organoid systems were successfully conducted through direct co-culture with microglia [51,52]. In addition, intestinal organoids have been actively applied to homeostasis regulation and disease mechanism studies by generating 3D multicellular organoids [53,54]. However, there are still challenges to be resolved to recapitulate the cellular diversity of human tissue, and there have been few reports on the function, maintenance, and disease modeling of organoids, including their populations of immune cells and other cells, especially in pulmonary AOs. To overcome this challenging, the optimization of culture media conditions in which diverse cells can maintain their phenotype and function will be the first step to mimicking the cellular diversity of human tissue. Mac-AOs papers reported so far have used AO media conditions for co-culture instead of optimizing appropriate co-culture conditions to maintain the two mixed cell types well [49,50]. Although, macrophages and epithelial cells were well maintained under the above conditions, optimization of co-culture medium conditions seems to be necessary to enable long-term culture and to maintain various functions and phenotypes of each cell. In brain organoids, organoid culture conditions with microglia were established by mixing basal medium of each cell type and adding cytokines necessary for the maintenance and maturation of each cell [51]. Another researcher focused on brain organoid basal media for co-culture and set up media conditions by adding cytokines to allow microglia to maintain function [52]. Based on the above, further study will be needed to optimize culture conditions that allow macrophages and epithelial cells to maintain their respective functions and phenotypes. Additionally, discovering factors that regulate the function and maintenance of both cells is also essential for the generation of organoids containing various types of cells.
hPSC-AOs: Maturity
Another limitation that needs to be addressed alongside cellular heterogeneity is ŌĆ£maturity,ŌĆØ or how similar the organoid is to human tissue. In fact, hPSC-derived AOs are not yet suitable for application in studies of age-related pulmonary fibrosis and infectious disease because they still more closely resemble fetal lung tissue than adult human lung tissue. Alysandratos et al. [55] showed distinct transcriptional and spatial differences between primary AT2 cells and hPSC-derived AT2 cells, with primary AT2 cells exhibiting more quiescent and mature characteristics, whereas hPSC-derived AT2 cells were more proliferative and immature, indicating an inverse relationship between the proliferative and maturation states of AT2 cells. The expression levels of AT1- and AT2-related genes were significantly lower in hPSC-derived AOs than in human primary cells, as were the expression levels of genes related to lamellar bodies, a measure of maturation. Recently, Lee et al. [56] evaluated the similarity between hPSC-derived LBOs and human lung tissue using a quantitative prediction system. LBOs exhibited 33.4% similarity to human lung tissue, whereas hESCs, DE cells, VAFECs, and human lung tissue showed similarities of 2.7%, 17.9%, 27.5%, and 100%, respectively [56]. The expression levels of AT2 cell-related genes, such as secreted frizzled-related protein 1 (SFPT1) and SFTPB, gradually increased following long-term cultivation. Although LBOs cultured for 56 to 59 days were used, their similarity to human lung tissue was still lower, and the cells showed immature fetal-like characteristics, such as low expression of the functional lung epithelial cell genes SFTPB and secretoglobin family 1A member 1 (SCGB1A1). These studies demonstrated the low similarity of current hPSC-derived organoids to human tissue and highlight the need for further studies to increase their maturity.
Recent studies offer some useful strategies to enhance AO maturity.
(1) Culture methods: Air-liquid interface culture (ALI) is a method for mimicking in vitro environment of gas exchange that is a unique feature of the lung and has been used as a useful platform to allow in vitro modeling of infection and environmental exposure. Recently, it has been reported that ALI culture up-regulates the expression level of SFTPC and AT2 maturation genes, while downregulating cell cycle-related transcripts compared to cultures in a 3D culture [57]. AT2 cells in ALI cultures were well maintained and upregulated their AT2 transcriptomic program at the single-cell level and highly expressed the key lineage markers SFTPC and NKX2.1 in protein level. In addition, ALI culture regulated tight junctions with an increase in transepithelial/transendothelial electrical resistance over time. These reports indicated that the ALI culture method was useful to maintain the AT2 phenotype while upregulating the expression of transcripts associated with AT2 maturation.
(2) Modulation of Wnt signaling: Jacob et al. [58] found that withdrawal of CHIR for 10 days significantly upregulated expression of SFTPC mRNA and other markers of AT2 cell maturation such as lysosomal associated membrane protein 3 (LAMP3) and solute carrier family 34 member 2 (SLC34A2). They also found that adding CHIR back after a 1-week following period further increased SFTPC proliferation and maintained its expression. In the culture conditions with CHIR modulation, the expression level of AT2-related genes such as SFTPC, SFTPB, and ABCA3 and additional AT2 maturation were increased. Several reports have shown that removing the glycogen synthase kinase 3 (GSK3) inhibitor at the initiation of the 3D cultures considerably have altered morphology and SFTPC marker expression [59]. Additionally, most of the cells in the CHIR withdrawal condition were cuboidal and there were significantly more lamellar bodies, indicating the maturity of the AT2 cells compared with CHIR treat condition as confirmed by transmission electron microscopy. These findings indicate that temporal regulation of Wnt signaling promotes AT2 cells maturation and proliferation.
As a basis for increasing organoid maturity, it is necessary to organize markers that can distinguish mature lung from fetal-like lung tissue. As described above, the major roles of AT2 cells include the synthesis and secretion of pulmonary surfactant for the maintenance of alveolar surface tension. Thus, most studies have identified SFTPC as a key marker distinguishing mature and functional AT2 cells from immature and non-AT2 cells during stepwise AEC differentiation from hPSCs. However, SFPTC has been reported to be expressed in both immature and mature AT2 cells and has also been identified as a bipotential and progenitor cell marker. Based on these findings, Sun et al. [29] suggested ABCA3 as a new marker of mature AT2 cells using an ABCA3:GFP and SFTPCtdTomato bifluorescent reporter hPSC line. SFTPCtdTomato+ cells were observed on day 19 of differentiation, whereas ABCA3:GFP+ cells emerged on day 22, in line with the known sequence of expression of SFTPC and ABCA3 in vivo during human lung development. In the sequence of human lung development, ABCA3 is detectable at approximately 28 weeks of gestation [60], and SFTPC is detectable at approximately 15 weeks of gestation. Therefore, ABCA3 is the most suitable gene for the differentiation of mature AT2 cells, together with other genes associated with lamellar bodies, as indicator markers of AT2 cell maturation (Figure 1). Though many studies have been conducted to identify markers that represent AT2 cell maturity, there are still few studies distinguishing AT1 cell-specific genes. The generation of more mature, advanced 3D multicellular AO culture systems will guide approaches for achieving a deeper understanding of human lung development and the pathogenesis of pulmonary disease.
Future Directions and Applications of hPSC-Derived AOs
As an alternative to mouse models and human primary cells, organoids have provided a useful platform for studying on pathogenesis and progression of pulmonary diseases, tissue homeostasis, and testing drug and environmental toxicity. However, organoids still have critical limitations to recapitulate the complex human lung tissue and AO research in various directions is lacking compared to the other organoids such as the brain and intestines. To address the AOs into the better research platform, two research directions will be needed.
As mentioned above, the incorporation of various cell types is essential to mimic the niche of human lung tissue and to understand the more detailed pathogenesis of pulmonary disease that occur through the interactions between multiple cells. Macrophages are the first defense system against various pathogens in alveoli and the key regulator of disease because they can control the disease progression according to their polarity [2,3]. Therefore, the generation of AOs containing macrophages is essential for disease modeling and provides a better understanding of pulmonary pathogenesis and drug testing. Although the generation of Mac-AOs using co-culture and forced aggregation methods have been reported [49,50], there are still many areas to be improved, such as regulating the polarity of macrophages within organoids and establishing long-term culture conditions. Furthermore, the incorporation of vascular cells remains an unexplored field. Vascular cells, which cover approximately 70% of the alveolar surface and control immune cell infiltration, are responsible for gas exchange, a unique characteristic of the lungs. Because vascular cells are key functional components in the lung, the generation of vascularized organoids will improve the complexity and function of organoids and facilitate fundamental studies on pulmonary development and diseases (Figure 1). Therefore, the establishment of advanced 3D multicellular AO culture systems will pave the way to unraveling many currently uncharacterized mechanisms related to pulmonary disease.
Another direction that organoids should consider along with generation of multicellular AOs is the ŌĆ£standardization of organoids.ŌĆØ Despite using the same cell line and the same differentiation method, hPSC-derived AOs typically exhibit heterogeneous sizes and morphologies, including folded and cystic structures [61]. Additionally, the regulation of AO apical-basal orientation still remains an unresolved area. These variations in generation of AOs make it difficult to expect consistent reactivity to drugs and environmental toxicants. Therefore, it is not suitable to be used as an evaluation platform that can completely replace experimental animals and there are limitations in providing an accurate mechanism of the pulmonary disease and the injury-repair systems. In order to standardize AOs, further research will be needed to identify novel factors that regulate the structure formation of organoids and establish media composition that can increase the maturity of organoids and maintain a constant cell composition ratio within AOs. The standardization of organoids through the discovery of novel methods that regulating structural formation and polarity of AOs will provide invaluable opportunities to study and understand all events that occur in the human lung.
Conclusion
In this review, we summarized the current methods of the generation of AOs from hPSCs and their useful applications in disease modeling and drug and toxicity evaluations. We also described the direction in which AO-related research should be oriented to achieve a more detailed representation of human lung tissue. hPSC-derived AOs are able to successfully mimic human alveoli in vitro , surpassing the limits of the low accessibility of human primary tissue and species differences, and have been used as useful platforms for pulmonary studies. Despite the great advancements that have been made in AO models, there is still work needed to improve the systems to fully realize the human lung alveolar environment, including issues related to cellular heterogeneity and maturity. The establishment of AOs containing other cells, especially macrophages and vascular cells, key components in lung function and pathogenesis, will begin to address the unknown factors that remain in the mechanisms of pulmonary disease. Furthermore, improvements in AO maturity will contribute to a more detailed understanding of human lung development and the pathogenesis of pulmonary diseases.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Print
Print Download Citation
Download Citation